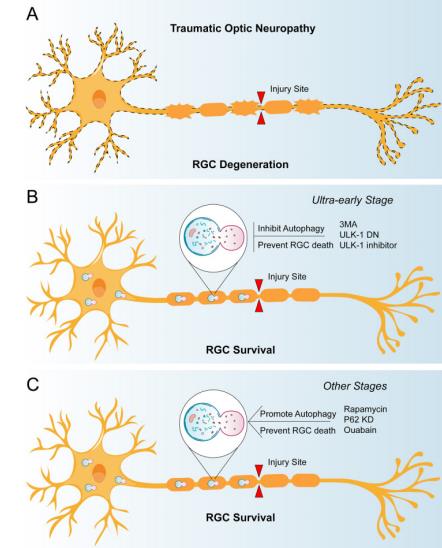
还有一些学者对视神经损伤后极早的时期进行研究,发现在该时期抑制自噬有利于减弱视神经损伤诱导的急性轴突变性。Kn?ferle等[19]发现在大鼠视神经挤压模型中,应用自噬抑制剂3-MA后90 min内,轴突变性显著延迟。作者推测对视网膜神经节细胞的机械损伤会通过激活钙通道诱导轴突内钙离子的快速增加,从而导致自噬体的二次积累,并参与轴突的退化过程,而自噬抑制剂3-MA减缓了这一过程。其中,钙离子可能通过钙调蛋白依赖的激酶β(CaMKKβ)激活腺苷酸激活蛋白激酶(AMPK),进一步抑制mTOR并激活自噬[26]。Koch等[18]在大鼠视神经挤压伤模型中观察到,视神经挤压伤后钙离子迅速流入轴突,诱导自噬启动,导致轴突解体。通过3-MA抑制自噬,可以显著减轻360 min内急性轴突变性。Vahsen团队发现过表达显性失活ULK1(dominate negative form of ULK1)可以抑制大鼠视神经挤压伤后的急性轴突变性并促进轴突再生[27-28]。同样,ULK1抑制剂SBI-0206965的应用也可以抑制大鼠视神经挤压伤后的急性轴突变性[27]。这些研究提示,在视神经损伤极早的时期,抑制自噬的激活有助于视神经的修复。值得注意的是,上述研究并未设置更长时间的观察节点,极早的时期过后自噬与轴突损伤的关系仍有待探索。
(A) In traumatic optic neuropathy, the optic nerve is damaged and the retinal ganglion cells degenerate progressively; (B) Before the ultra-early stage of traumatic optic neuropathy, the use of 3-MA, ULK-1 DN or ULK-1 inhibitor can inhibit autophagy after optic nerve injury, alleviate acute axonal injury, and be conducive to the survival of retinal ganglion cells; (C) After the ultra-early stage of traumatic optic neuropathy, the survival rate of retinal ganglion cells increased after the application of autophagy activators such as rapamycin and p62 siRNA.
1. 国家自然科学基金(82171404);中山大学科研领军人才培育计划项目(22yklj04);科研启动经费(高水平医院建设);国家重点研发计划 (2022YFF1202901)。 This work was supported by National Natural Science Foundation of China(82171404); Sun Yat-sen University Scientific
Research Leading Talents Cultivation Program(22yklj04); Scientific research start-up funds (high-level hospital construction); National key research and development
program(2022YFF1202901)
参考文献
1. Ribas VT, Vahsen BF, Tatenhorst L, et al. AAV-mediated inhibition of
ULK1 promotes axonal regeneration in the central nervous system in
vitro and in vivo[ J]. Cell Death Dis, 2021, 12(2): 213.
2. Vahsen BF, Ribas VT, Sundermeyer J, et al. Inhibition of the autophagic
protein ULK1 attenuates axonal degeneration in vitro and in vivo,
enhances translation, and modulates splicing[ J]. Cell Death Differ,
2020, 27(10): 2810-2827.
3. Hu YX, Han XS, Jing Q. Ca(2+) ion and autophagy[ J]. Adv Exp Med
Biol, 2019, 1206: 151-166.
4. Dehay B, Bové J, Rodríguez-Muela N, et al. Pathogenic lysosomal
depletion in Parkinson's disease[ J]. J Neurosci, 2010, 30(37): 12535-
12544.
5. Mázala-de-Oliveira T, de Figueiredo CS, de Rezende Corrêa
G, et al. Ouabain-Na+/K+-ATPase signaling regulates retinal
neuroinflammation and ROS production preventing neuronal death by
an autophagy-dependent mechanism following optic nerve axotomy in
vitro[ J]. Neurochem Res, 2022, 47(3): 723-738.
6. Wei J, Ma L S, Liu D J, et al. Melatonin regulates traumatic optic
neuropathy via targeting autophagy[ J]. Eur Rev Med Pharmacol Sci,
2017, 21(21): 4946-4951.
7. Wen YT, Zhang JR, Kapupara K, et al. mTORC2 activation protects
retinal ganglion cells via Akt signaling after autophagy induction in
traumatic optic nerve injury[ J]. Exp Mol Med, 2019, 51(8): 1-11.
8. Rodríguez-Muela N, Germain F, Mari?o G, et al. Autophagy promotes
survival of retinal ganglion cells after optic nerve axotomy in mice[ J].
Cell Death Differ, 2012, 19(1): 162-169.
9. Koch JC, Lingor P. The role of autophagy in axonal degeneration of the
optic nerve[ J]. Exp Eye Res, 2016, 144: 81-89.
10. Kn?ferle J, Koch J C, Ostendorf T, et al. Mechanisms of acute axonal
degeneration in the optic nerve in vivo[ J]. Proc Natl Acad Sci U S A,
2010, 107(13): 6064-6069.
11. Koch JC, Kn?ferle J, T?nges L, et al. Acute axonal degeneration in
vivo is attenuated by inhibition of autophagy in a calcium-dependent
manner[ J]. Autophagy, 2010, 6(5): 658-659.
12. Kim SH, Munemasa Y, Kwong JM, et al. Activation of autophagy in
retinal ganglion cells[ J]. J Neurosci Res, 2008, 86(13): 2943-2951.
13. Klionsky DJ, Abdel-Aziz AK, Abdelfatah S, et al. Guidelines for the use
and interpretation of assays for monitoring autophagy(4th edition)[ J].
Autophagy, 2021, 17(1): 1-382.
14. Galluzzi L, Green DR . Autophagy-independent functions of the
autophagy machinery[ J]. Cell, 2019, 177(7): 1682-1699.
15. Wong PM, Puente C, Ganley IG, et al. The ULK1 complex: sensing
nutrient signals for autophagy activation[ J]. Autophagy, 2013, 9(2):
124-137.
16. Ma Q, Long S, Gan Z, et al. Transcriptional and post-transcriptional
regulation of autophagy[ J]. Cells, 2022, 11(3): 441.
17. Boya P, Reggiori F, Codogno P. Emerging regulation and functions of
autophagy[ J]. Nat Cell Biol, 2013, 15(7): 713-720.
18. Shen S, Kepp O, Kroemer G. The end of autophagic cell death? [ J].
Autophagy, 2012, 8(1): 1-3.
19. Marshall RS, Vierstra RD. Autophagy: the master of bulk and selective
recycling[ J]. Annu Rev Plant Biol, 2018, 69: 173-208.
20. Boya P. Why autophagy is good for retinal ganglion cells? [ J]. Eye
(Lond), 2017, 31(2): 185-190.
21. Stavoe AKH, Holzbaur ELF. Autophagy in neurons[ J]. Annu Rev Cell
Dev Biol, 2019, 35: 477-500.
22. Li HY, Ruan YW, Ren CR, et al. Mechanisms of secondary degeneration
after partial optic nerve transection[ J]. Neural Regen Res, 2014, 9(6):
565-574.
23. Bastakis GG, Ktena N, Karagogeos D, et al. Models and treatments for
traumatic optic neuropathy and demyelinating optic neuritis[ J]. Dev
Neurobiol, 2019, 79(8): 819-836.
24. Bacorn C, Morisada MV, Dedhia RD, et al. Traumatic optic neuropathy
management: a survey assessment of current practice patterns[ J]. J
Emerg Trauma Shock, 2021, 14(3): 136-142.
25. Xie D, Yu H, Ju J, et al. The outcome of endoscopic optic nerve
decompression for bilateral traumatic optic neuropathy[ J]. J Craniofac
Surg, 2017, 28(4): 1024-1026.
26. Wladis EJ, Aakalu V K, Sobel R K, et al. Interventions for indirect
traumatic optic neuropathy: a report by the American academy of
ophthalmology[ J]. Ophthalmology, 2021, 128(6): 928-937.
27. Lee V, Ford RL, Xing W, et al. Surveillance of traumatic optic
neuropathy in the UK[ J]. Eye (Lond), 2010, 24(2): 240-250.
28. Karimi S, Arabi A, Ansari I, et al. A systematic literature review on
traumatic optic neuropathy[ J]. J Ophthalmol, 2021, 2021: 5553885.
'%20fill='white'%20fill-opacity='0.01'/%3e%3cmask%20id='mask0_3477_29692'%20style='mask-type:luminance'%20maskUnits='userSpaceOnUse'%20x='0'%20y='0'%20width='16'%20height='16'%3e%3crect%20id='&%23232;&%23146;&%23153;&%23231;&%23137;&%23136;_2'%20x='16'%20width='16'%20height='16'%20transform='rotate(90%2016%200)'%20fill='white'/%3e%3c/mask%3e%3cg%20mask='url(%23mask0_3477_29692)'%3e%3cpath%20id='&%23232;&%23183;&%23175;&%23229;&%23190;&%23132;'%20d='M14%205L8%2011L2%205'%20stroke='%23333333'%20stroke-width='1.5'%20stroke-linecap='round'%20stroke-linejoin='round'/%3e%3c/g%3e%3c/g%3e%3c/svg%3e)