小胶质细胞调控视网膜类器官中成熟及血管化的研究进展
'%20fill='white'%20fill-opacity='0.01'/%3e%3cmask%20id='mask0_3477_29692'%20style='mask-type:luminance'%20maskUnits='userSpaceOnUse'%20x='0'%20y='0'%20width='16'%20height='16'%3e%3crect%20id='&%23232;&%23146;&%23153;&%23231;&%23137;&%23136;_2'%20x='16'%20width='16'%20height='16'%20transform='rotate(90%2016%200)'%20fill='white'/%3e%3c/mask%3e%3cg%20mask='url(%23mask0_3477_29692)'%3e%3cpath%20id='&%23232;&%23183;&%23175;&%23229;&%23190;&%23132;'%20d='M14%205L8%2011L2%205'%20stroke='%23333333'%20stroke-width='1.5'%20stroke-linecap='round'%20stroke-linejoin='round'/%3e%3c/g%3e%3c/g%3e%3c/svg%3e)
关键词
摘要
诱导多能干细胞衍生的视网膜类器官(retinal organoids, ROs)作为重要的体外模型,已成为研究视网膜发育规律和构建体外疾病模型的重要工具。然而,传统ROs尚缺乏小胶质细胞等免疫成分,限制了其在模拟生理状态下神经-免疫细胞互作及病理过程中神经炎症与免疫反应中的应用。本文综述了近年来将小胶质细胞整合到ROs中的研究进展,梳理了外源性共培养、内源性自组装以及基于3D打印与微流控器官芯片的构建策略,比较了不同的ROs整合小胶质细胞方案的特征。深入阐述了小胶质细胞在ROs发育中的调控机制,其通过突触修剪促进神经环路的成熟,通过分泌细胞因子引导内皮细胞迁移从而诱导类器官血管网络形成。并探讨了复合模型在模拟眼科相关疾病中的应用。同时,总结了小胶质细胞在介导神经炎症,清除光感受器碎片及在肿瘤免疫微环境中的具体功能,为深入理解眼部疾病的致病机制以及为小胶质细胞在视网膜发育及视网膜神经退变疾病过程中的作用提供了新的思路和见解。最后,文章展望了基于微流控和器官芯片技术的模型构建方向,指出免疫功能完善的ROs为视网膜疾病机制解析和药物筛选提供更精准的研究平台。
全文
文章亮点
1. 关键发现
• 本文系统综述了小胶质细胞在视网膜类器官的神经环路成熟、血管网络形成及神经炎症响应方面的关键调控作用。
2. 已知与发现
• 传统视网膜类器官缺乏小胶质细胞和血管系统,限制了其在模拟神经炎症和复杂病理过程中的应用。小胶质细胞的整合优化了类器官的免疫功能,并可促进视网膜及脑类器官的成熟与血管化。
3. 意义与改变
• 本综述为我们深入理解眼部疾病的致病机制以及为小胶质细胞在视网膜发育及视网膜神经退变疾病过程中的作用提供了新的思路和见解。
视网膜类器官(retinal organoids, ROs)是利用诱导多能干细胞(induced pluripotent stem cells,iPSCs)在体外诱导分化形成具有多种视网膜神经细胞的三维组织结构,其在结构和功能上能较好模拟人视网膜的特征[1-2]。与传统的二维细胞培养相比,ROs具有类视网膜层状结构以及包含人视网膜中的所有主要细胞类型,包括感光细胞、视网膜神经节细胞(retinal ganglion cells, RGCs)、穆勒细胞(Müller cells)、双极细胞和水平细胞等[1, 3-4]。此外,ROs在基因表达模式、mRNA可变剪接和电生理功能上与人类视网膜高度相似,能在体外近似模拟人类疾病的病理生理动态演变过程[1, 5]。因此,ROs在视网膜疾病发病机制研究及药物筛选中具有广阔的应用前景[6-7]。然而,目前常用的ROs仍存在诸如感光细胞成熟度低、存活和稳定性不高以及免疫功能缺如等不足,而这些缺陷与ROs中缺失血管和免疫细胞密切相关[8-11]。
小胶质细胞是视网膜中主要的免疫细胞,具有免疫监测与应答、吞噬、调节炎症反应和化学环境、促进神经元和网络成熟、突触修剪、营养支持和调节血管生成等作用[11-14]。在病理状态下,持续或异常激活的小胶质细胞会释放神经毒性因子,降低吞噬效率和驱动神经炎症,从而加速神经元损伤和疾病进展[14]。由于ROs和小胶质细胞在胚胎起源上的根本差异,常用的ROs模型普遍缺乏小胶质细胞等免疫细胞成分[11, 15]。而缺乏小胶质细胞的ROs无法模拟神经元-胶质细胞之间复杂且动态的相互作用,从而限制了ROs在年龄相关性黄斑变性(age-related macular degeneration, AMD)、糖尿病性视网膜病变(diabetic retinopathy, DR)和青光眼等涉及神经炎症的退行性疾病中的研究[11, 16-18]。
ROs模型虽展现出强大的应用潜力,但其缺乏内源性小胶质细胞成分和功能性血管化系统,这极大制约了该模型在疾病模拟和机制解析中的应用。因此,小胶质细胞与ROs的有效整合正成为ROs研究领域的焦点。为了利用ROs复合模型来深入阐释疾病生理与病理过程,并作为平台评估药物开发及潜在治疗方法,本文系统总结当前将小胶质细胞引入ROs的各种策略,以及其在ROs成熟及血管化中的作用,以期为该领域的未来发展和相关研究提供新的见解。
1. 视网膜类器官“胶质化”的方法
ROs“胶质化”,即ROs与小胶质细胞的整合,可通过共培养(co-culture)和自组织(self-organization)两种方式获得[11, 19-20]。目前主要通过分别分化ROs和小胶质细胞,再利用共培养的方式将小胶质细胞整合到ROs的方法来获得具有小胶质细胞的ROs。
1.1 小胶质细胞与ROs共培养获得“胶质化”ROs
共培养是目前最主要且有效地获得“胶质化”ROs的方式[11, 19, 21](图1)。其基本方法是将iPSC来源的ROs和小胶质细胞样(induced micoglia-like cell, iMG)细胞进行共同培养[11]。其中小胶质细胞的来源主要分为3种:1)由iPSC直接分化为iMG;2)由iPSC分化为小胶质前体细胞(microglial precursor cell, preMG);3)由iPSC分化为巨噬细胞祖细胞[22-24]。根据不同的研究目的,iMG可在ROs分化的不同时期引入,iMG从圆形或阿米巴样形态转变为具有复杂分支的静息态小胶质细胞形态,并能够主动迁移并整合到类器官内部,定植于与体内视网膜小胶质细胞相应的解剖位置[11, 21, 25-26]。通过共培养的方式,iMG能够与ROs中的神经元和胶质细胞进行互作,整合后的iMG一方面会表现出形态的成熟,另一方面具有显著增强的功能和细胞器成熟度,包括K+通道表达增强和线粒体、内质网等细胞器的成熟度增加[11, 22, 25]。同时,ROs微环境中的非接触性信号反过来可以促进iMG的功能成熟[25, 27]。
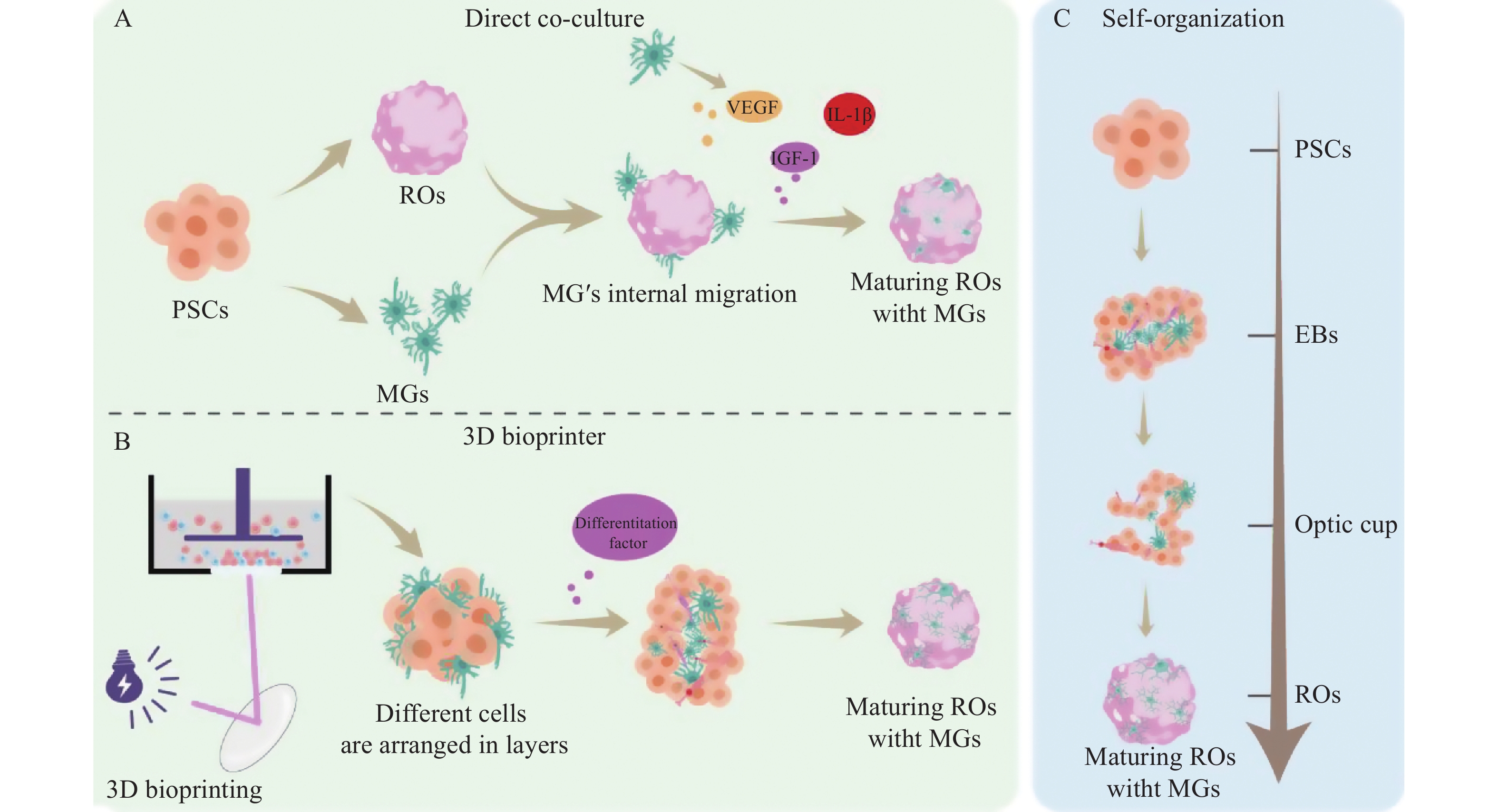
PSCs: 多能干细胞;EBs: 类胚体;ROs: 视网膜类器官。
PSCs: Pluripotent stem cells; EBs: Embryoid bodies; ROs: Retinal organoids.
在ROs与iMG的融合中,3D生物打印是未来的主流方法。3D生物打印技术是将活细胞整合到特殊水凝胶基质的生物墨水中,然后通过打印机精确地构建出三维结构[28-29](图1B)。该技术也可以应用于小胶质细胞的引入。为了更好地模拟视网膜丛状层和细胞外基质特性,研究人员使用由明胶和透明质酸组成的特定硬度的水凝胶作为生物墨水,已成功利用3D数字光处理(digital light processing, DLP)生物打印技术构建了由内皮细胞和小胶质细胞组成的共培养模型,以模拟DR的病理微环境[30-31]。这种3D生物打印模型克服了传统2D培养无法模拟复杂细胞间相互作用和细胞外基质特性的局限性[31]。微流控(microfluidics)是在微米级别的空间中精确操控微量液体的技术,能高度模拟体内的动态血流动力学环境[32]。而器官芯片(organ-on-a-Chip)通过将微流控操作系统和生物培养分析技术集成在芯片上,来培养活细胞以模拟人体的生理环境[33]。利用3D打印和微流控系统生成的管状类脑器官通过流动培养基将诱导的iMG自然迁移至类器官中,这种新型的引入方式避免了注射带给类器官的损伤,并使得类脑器官中的细胞因子响应更强,且神经元与胶质细胞的互作更接近体内的生理状态[34]。利用微流控和器官芯片技术可以为培养ROs提供持续的营养供应,并带走代谢产物,其提供的模拟生理状态的血管功能可以促进ROs的成熟和长期维持培养,在模拟人类疾病和开发治疗策略方面具有潜在应用价值[35]。
1.2 自主分化方法获得“胶质化”ROs
在特定培养条件下,iPSCs能够直接分化形成包含小胶质细胞在内的所有类型视网膜细胞,并形成与活体视网膜类似的三维层状结构[36-38]。与共培养模型相比,自组织模型不需要将小胶质细胞分开单独分化,更接近视网膜生理发育过程,且能够为小胶质细胞的发育提供微环境(图1C)。目前小胶质细胞在脑类器官中的自主分化的技术更为成熟,可以通过调控骨形态发生蛋白信号,将iPSCs分化为中胚层来源的祖细胞,并且通过干预神经诱导信号将中胚层细胞分化为小胶质细胞[39]。ROs的自组织分化方案借鉴脑类器官的分化方案。iPSCs经过类器官分化的初始悬浮培养阶段形成胚胎体[40]。胚胎体自发形成神经外胚层,并进一步发育为视杯[40-41]。IBA1+小胶质细胞样细胞在培养的第3~4周出现,主要富集于非色素性的3D囊泡区[40]。囊泡区表达间充质和上皮标志物,为小胶质细胞的驻留提供了微环境,该区域小胶质细胞的表型和发育轨迹模拟了体内小胶质细胞的早期发育[20]。但类器官内的小胶质细胞比例仍是高度可变的,仍需进一步研究来生成比例稳定的类器官模型[42]。在脑类器官和小胶质细胞的融合中,共培养组的iMG细胞呈现聚集样分布,细胞成熟度更高,而经过自主分化的iMG分布更均匀且形态接近稳态[43]。
因此,研究人员在选择小胶质细胞整合策略时,必须权衡自组织分化与共培养方法的优势和局限性。自组织分化更贴合生理发育时间轴,能确保小胶质细胞在结构上分布均匀,并维持接近静息形态和较低的基线激活水平,使其成为研究正常发育和慢性炎症的研究模型[20, 43]。相比之下,共培养方法虽然能够更灵活地控制小胶质细胞的比例,但其细胞倾向于呈聚集样分布,这使其更适用于研究急性或高密度小胶质细胞参与的病理过程[43-44]。最终整合方法的选择应严格取决于特定的研究目标和所需的细胞表型状态。
2. 小胶质细胞对视网膜类器官生理功能的影响
2.1 小胶质细胞对类器官成熟分化的影响
小胶质细胞在视网膜的发育和细胞成熟中扮演着不可或缺的角色,其通过独特的吞噬功能优化神经环路[45-46]。在视网膜发育过程中,神经元会产生大量的突触连接。小胶质细胞通过其高度发达的突起,持续监控局部微环境,并选择性地吞噬和修剪多余或功能不佳的突触,从而优化神经环路[47-48]。小胶质细胞突触修剪可以通过吞噬途径和非吞噬途径来进行。吞噬途径中,小胶质细胞通过吞噬游离的突触后蛋白,延缓双极细胞异常突起的形成,从而改善视网膜神经环路[49]。非吞噬途径中,小胶质细胞通过补体受体识别被补体蛋白标记的突触,抑制异常神经突起的生长从而延缓视觉功能恶化[49-53]。无论是通过吞噬游离蛋白改善视网膜重构,还是通过补体介导的途径抑制异常突起生长,小胶质细胞的修剪作用对于类器官中神经元的发育和结构完善都起到了不可或缺的促进作用[20, 45-46, 54](图2A)。
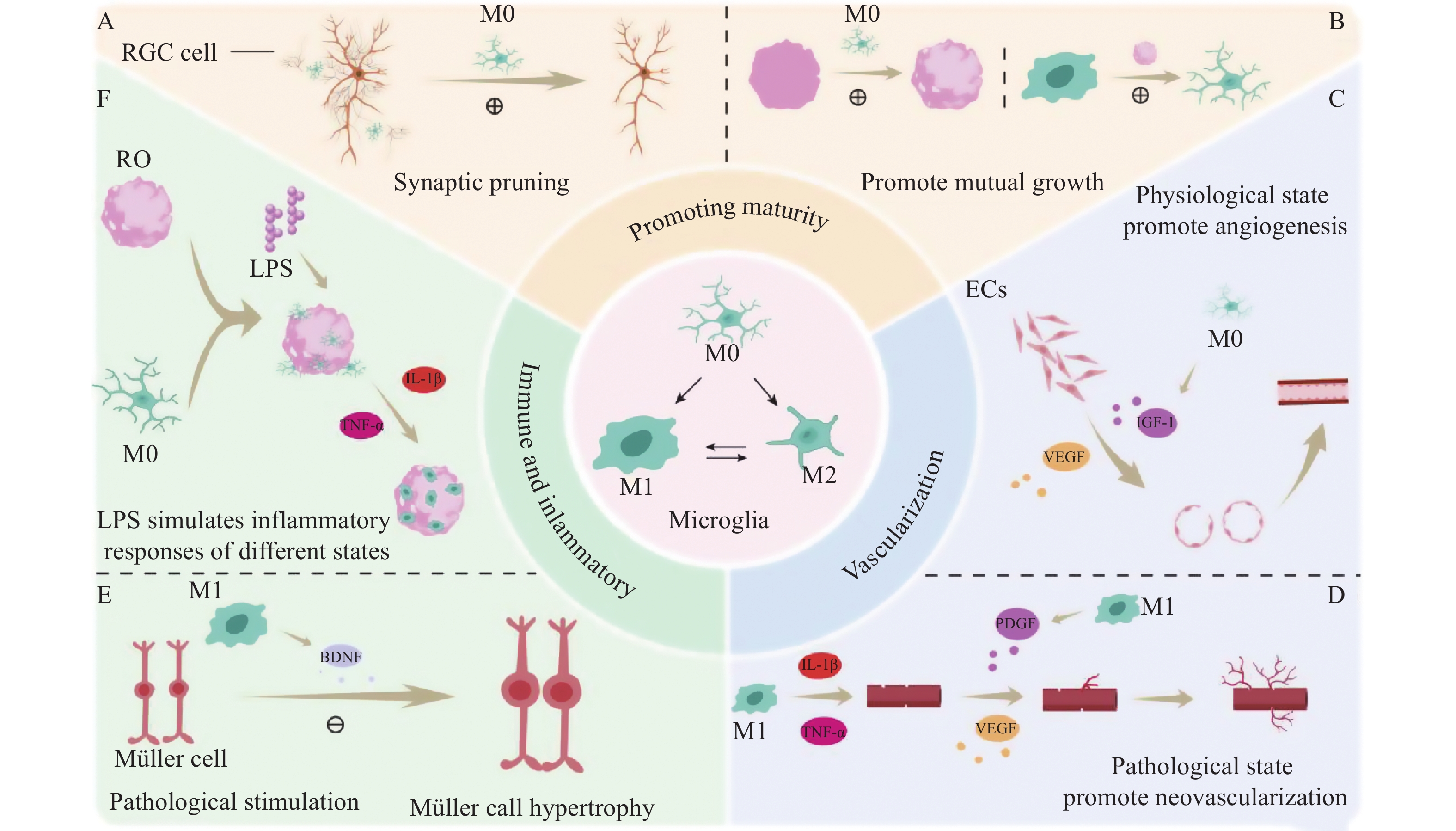
ROs: 视网膜类器官;LPS: 脂多糖;ECs: 内皮细胞;VEGF: 血管内皮生长因子;PDGF: 血小板衍生生长因子;RGCs: 视网膜神经节细胞。
ROs: Retinal organoids; LPS: Lipopolysaccharide; ECs: Endothelial cells; VEGF: Vascular endothelial growth factor; PDGF: Platelet-derived growth factor; RGCs: Retinal ganglion cells.
小胶质细胞可以通过多种方式促进ROs的成熟。首先,小胶质细胞通过分泌细胞因子调控视网膜细胞分化来维持视网膜的内环境稳定[46]。整合iMGs能够促进类器官的整体成熟,这一点在脑类器官模型中已得到证实,并且在ROs中也存在类似的功能[11]。在ROs中,小胶质细胞通过分泌抗炎因子或者细胞间信号传导间接稳定内皮细胞的表达,从而参与血-视网膜屏障(blood-retinal barrier, BRB)的稳态维持[21, 24]。其次,小胶质细胞在ROs中迁移,通过与巨噬细胞相互作用调节视网膜屏障稳态及维持血-视网膜屏障结构的完整性[21, 24]。在ROs和小胶质细胞进行整合时,iMG由靠近神经节细胞的视杯腔内逐渐朝视网膜外丛状层延伸其突起,iMG可以通过吞噬作用调节神经节细胞的数量来促进ROs的成熟性和功能完整性[22]。最后,小胶质细胞通过共同参与形成三维神经血管单元结构,对类器官的成熟及复杂结构的完善发挥积极作用[55]。研究者将iMG整合到血管化的ROs得到了含有iMG的血管化ROs(vascularized retinal organoids, vROs)的模型,并对其进行转录组学分析,发现相较于血管化的视网膜类器官本身,含有iMG的vROs中差异表达基因显著富集于免疫反应和血管生成通路[55]。这提示小胶质细胞可以促进ROs的血管化以及免疫功能的完善。
此外,小胶质细胞与类器官的成熟是相互促进的, 类器官的成熟也为小胶质细胞的发育与功能的执行起到促进作用[23]。共培养时类器官内的微环境为小胶质细胞的发育成熟提供了基质环境[56]。在与类器官共培养后,共培养环境提升了小胶质细胞溶酶体和线粒体等关键细胞器的成熟度,小胶质细胞的吞噬能力显著增强,这种跨细胞类型的相互作用,反过来又增强了小胶质细胞对类器官的积极影响[25](图2B)。vROs通过共培养血管类器官(vascular organoids, VOs)和ROs实现了iMG的整合,形成的血管微环境进一步促进了iMG的迁移和驻留[55]。
2.2 小胶质细胞对类器官血管化的影响
小胶质细胞在视网膜血管系统发育与稳态中发挥着关键调控作用[57]。小胶质细胞可以通过分泌多种血管生成因子,如血管内皮生长因子(vascular endothelial growth factor, VEGF)和胰岛素样生长因子1(insulin-like growth factor-1, IGF-1),调节内皮细胞的增殖和迁移,从而促进血管网络的形成[58]。与类脑器官联合培养后,小胶质细胞除了迁移至神经元区域,修剪神经元突触,来维持神经血管单元的稳态外,iMG还可迁移至血管内皮细胞周围,并且通过分泌细胞因子调节内皮紧密连接或星形胶质细胞活性[59- 60]。此外,VOs也可以反过来促进小胶质细胞分化[59, 61]。而分化后的iMG表达多种成熟小胶质细胞标记物,并具备免疫监视能力,可通过吞噬清除血管周围病原体或凋亡碎片,维持血管完整性[60, 62](图2C)。因此,小胶质细胞与VOs能相互作用,促进彼此的成熟。
在病理条件下,缺氧和高血糖等因素会激活小胶质细胞,使其转变为促炎、促新生血管生成的M1极化状态[63- 64]。这些活化的M1型小胶质细胞积累在新生血管簇周围,并通过分泌大量促血管生成因子和促炎因子,如血小板衍生生长因子(platelet-derived growth factor, PDGF)、白细胞介素-1β(interleukin- 1β, IL-1β)和肿瘤坏死因子-α (tumor necrosis factor-α, TNF-α),来诱导病理性血管生成和紧密连接蛋白的破坏,同时调节内皮细胞的状态[24, 65]。这种异常的新生血管伴随着BRB的破坏,导致不可逆的视网膜损伤[24] (图2D)。最新研究利用整合了小胶质细胞的vROs模型来模拟DR,并证实小胶质细胞在调控血管结构中具有关键作用[66]。在高糖环境中,小胶质细胞表现出激活状态,形态转变为阿米巴样的M1型小胶质细胞,并显示出更高的IBA1蛋白表达[31]。活化的小胶质细胞能够促进视网膜微血管内皮细胞的迁移和血管生成,同时减少紧密连接蛋白的产生,表明小胶质细胞与内皮细胞之间存在动态相互作用[58]。转录组学研究发现,小胶质细胞基因表达与血管发育、细胞迁移和对缺氧反应等途径显著相关,凸显了小胶质细胞在调节血管结构和应对病理生理变化中的潜在关键作用[31]。
2.3 小胶质细胞在类器官中的炎症响应
小胶质细胞是视网膜的常驻免疫细胞,它的整合赋予类器官的免疫响应能力[11]。在正常生理条件下,小胶质细胞对维持视网膜的稳态、清除凋亡细胞或重塑突触发挥重要作用;然而在病理状态下,iMG会被迅速激活为M1或M2型反应性表型[23, 45, 67-68]。在持续激活的状态下,小胶质细胞会分泌多种神经毒性因子,导致视网膜神经元继发性死亡,从而加速神经退行性病变[45, 69]。
在脑类器官模型中,小胶质细胞可与其他胶质细胞进行互作,并通过激活信号通路和释放炎症因子来诱导星形胶质细胞的病理性胶质化,因此建立整合小胶质细胞的类器官共培养模型有助于进一步理解疾病的病理过程[70-72](图2E)。整合携带疾病风险基因的iMG与类器官的联合培养已成为研究神经退行性疾病机制的有力工具,为阐明小胶质细胞在阿尔茨海默病(Alzheimer's disease, AD)和帕金森病(Parkinson's disease, PD)病理中的作用提供了关键证据[73-74]。研究者们将携带疾病风险基因的小胶质细胞与AD模型的类器官共培养,发现携带疾病风险基因的小胶质细胞对病理沉积物的清除能力显著下降,进而导致病理沉积物的蓄积[75]。此外,将帕金森病患者来源的iMG整合至类脑器官后,可导致类脑器官内PD病理蛋白的出现,这直接证实了病变的小胶质细胞在驱动PD病理中的核心作用[76-77]。
同样,ROs在解析人类视网膜发育机制及模拟复杂病理过程中发挥重要作用。ROs能够以高度拟人化的时空模式形成视网膜的层级结构与多细胞功能单位,从而实现对视网膜相关疾病的精准建模。通过患者来源的hiPSCs(如携带USH2A或PDE6B突变),能够再现视网膜色素变性(retinitis pigmentosa, RP)的核心病理特征,包括光感受器的进行性退化、早期神经上皮发育异常、细胞凋亡过程及光转导通路的异常[78-79]。并且通过共培养技术将小胶质细胞成功整合至RP类器官模型中,证实了布洛芬等非甾体抗炎药(non-steroidal anti-inflammatory durgs, NSAIDs)在调控小胶质细胞活性中的治疗潜力,为通过调节小胶质细胞活性来延缓视力丧失的精准治疗提供了关键的体外研究平台[22]。此外,多位研究者利用分化成熟的ROs构建了干性AMD模型,成功再现了视网膜色素上皮(retinal pigment epithelium, RPE)和光感受器的凋亡病理,并筛选出二甲双胍等潜在保护药物,但目前该模型缺乏血管系统与免疫反应[80-81]。de Lemos等[82]在2024年利用ROs成功构建了早期DR模型,并证实了Müller细胞在DR发病早期可通过调节炎症反应参与疾病的发生[83]。同样,研究者通过人胚胎干细胞(human embryonic stem cells, hESCs)诱导可获得人源视网膜母细胞瘤(retinoblastoma, RB)类器官模型,借助该模型研究者探讨了人类RB的致癌起源,并评估了化疗药物的毒性和抗肿瘤效果及侵袭性等肿瘤特性[82]。2025年Xu等[84]构建了包含功能性小胶质细胞的RB类器官模型,成功模拟了免疫细胞对3D肿瘤组织的靶向识别与清除过程。工程化的小胶质细胞在ROs内表现出特异性的抗肿瘤活性,这一模型的建立为解析小胶质细胞在视网膜病理状态下的表型转化及其在视网膜恶性肿瘤中的治疗提供了依据[85]。综上所述,ROs模型的潜力有待进一步探索,未来研究需要专注于优化小胶质细胞的功能性整合和开发靶向神经炎症的治疗策略[85-86]。
在共培养模型中,iMG能够持续发挥其常驻免疫细胞的功能,即通过伸缩的突起对视网膜微环境进行实时监测[46]。当模型暴露于病理刺激物时,iMG能迅速激活炎症基因的表达,并释放一系列促炎细胞因子放大炎症信号[11, 25]。通过改变LPS的水平和处理时间,研究人员可以模拟急性和慢性炎症状态,从而系统性地研究小胶质细胞在不同病理阶段的动态变化及其对其他视网膜细胞的影响,为探究神经炎症在视网膜疾病中的作用提供了研究平台[25](图2F)。
3. 总结与展望
小胶质细胞在视网膜的发育、稳态维持及应对损伤中发挥着至关重要的作用。为弥补传统ROs缺乏免疫细胞和功能性血管系统的缺陷,研究人员已成功构建了整合小胶质细胞的ROs体外模型,并在血管化方面取得了初步进展。然而,现有模型在模拟体内复杂生理功能方面仍存在不足:挑战在于如何构建具备完整功能的血管化网络和反映体内真实状态的免疫微环境。目前,类器官模型中的“血管化”大多采用无细胞的灌注通道,这远不能复制体内高度复杂的血管网络结构,并且导入类器官内的小胶质细胞比例及存活状态仍需优化。其次,现有ROs在结构和功能上尚未完全达到成年视网膜的成熟水平;长期培养中,营养和氧气限制常导致视网膜神经上皮层细胞数量减少,因此实现长期的、完全成熟的培养条件亟待优化。
未来研究应致力于将小胶质细胞-ROs模型与微流控技术和器官芯片系统进行深度结合。通过这些技术,实现对微环境参数(如氧气、营养物质和剪切力梯度)的精准动态调控,从而构建出更具生理学相关性的复杂血管组织-免疫系统复合模型。这种高度整合、功能化的平台将成为强大的疾病研究工具,不仅能够用于深入解析小胶质细胞功能在视网膜病理中的作用机制,还可高效筛选和评估靶向神经炎症的新型药物治疗策略。
声明
在本作品的准备过程中,作者使用了DeepSeek V3进行语言润色和部分文献调研,使用此工具后,作者根据需要对内容进行了审查和编辑,并对出版物的内容承担全部责任。
利益冲突
所有作者均声明不存在利益冲突。
开放获取声明
本文适用于知识共享许可协议(Creative Commons),允许第三方用户按照署名(BY)-非商业性使用(NC)-禁止演绎(ND)(CC BY-NC-ND)的方式共享,即允许第三方对本刊发表的文章进行复制、发行、展览、表演、放映、广播或通过信息网络向公众传播,但在这些过程中必须保留作者署名、仅限于非商业性目的、不得进行演绎创作。
基金
1. 科技部国家重点研发计划(2022YFF1202901);国家自然科学基金(82171404);中央高校基本科研业务费专项资金(22yklj04);广东省自然科学基金项目(2023A1515011529);广州市科技计划项目(2023A03J0181,2024A04J6481);中山大学科研启动基金(人才基金〔2020〕18);眼病防治全国重点实验室课题基金(2025QZSPT34)。This work was supported by the Key Research and Development Program of the Ministry of Science and Technology of the People's Republic of China (2022YFF1202901), the National Natural Science Foundation of China(82171404), the Fundamental Research Funds for the Central Universities (22yklj04), the Natural Science Foundation of Guangdong Province of China (2023A1515011529), the Science and Technology Planning Project of Guangzhou City(2023A03J0181, 2024A04J6481), the Research Start-up Founds of Sun Yat-sen University (Funded Talent 〔2020〕18), and the Research Funds of the State Key Laboratory of Ophthalmology (20250ZSPT34).
参考文献
Xu J, Jin ZB. Monitoring effect of microglia on retinal and nervous system diseases[J]. Chin J Exp Ophthalmol, 2022, 40(8): 758-764. DOI: 10.3760/cma.j.cn115989-20200918-00656.
Huo YJ, Chen J, Zhang AM, et al. The role of complement system in mental diseases[J]. J Cent South Univ (Med Sci), 2023, 48(10): 1539-1545. DOI: 10.11817/j.issn.1672-7347.2023.230109.