


HIGHTLIGHTS
1.Critical Discoveries and Outcomes
This review synthesizes evidence from population studies and experimental research to elucidate the interaction of meibomian gland obstruction and dropout in the pathogenesis of meibomian gland dysfunction.
2.Methodological Innovations
This review delves into the intricate interaction between ductal obstruction and glandular dropout within the meibomian gland, based on the structural and functional distinction of ductal and acinar epithelia.
3. Prospective Applications and Future Directions
The potential pathological mechanisms summarized in this study offer valuable insights that could inform the early diagnosis and intervention of meibomian gland dysfunction.
The pathogenesis of MGD remains elusive. Previous hypothesis supported that the initial step of MGD involved gland obstruction due to hyperkeratinization of the ductal epithelium, leading to duct dilation and chronic intraglandular high pressure, ultimately causing secondary glandular dropout [3, 11]. However, this theory failed to fully account for certain observations in MGD patients, such as those with age-related MGD, who displayed significant glandular dropout with minimal evidence of obstruction [12]. It was known that the risk factors for MGD related to meibum metabolism disorder or damage to the MG ductal epithelial cells include chronic inflammatory infiltration and fluctuations in sex hormones [13-15]. These factors can lead to ducts hyperkeratinization and imbalance in meibum lipid composition, resulting in intraductal meibum stagnation, thereby inducing MG blockage, such prolonged blockage further increased intraductal pressure, ultimately induced functional acinar atrophy [16-17]. However, under the influence of aging and benzalkonium (BAK) chloride exposure [18-19], significant loss of progenitor cells and acinar cells can occur independently without apparent MG obstruction. Characterizing MG dropout solely as a progressive result of MG ductal obstruction overlooks the early diagnostic potential of glandular dropout, which exhibits high sensitivity (96.7%) and specificity (85%) in identifying MGD [20]. Notably, the loss of MG acini also affected meibocyte differentiation and meibum metabolism, increasing the risk of MG obstruction [21-22]. These findings indicate that glandular dropout and ductal obstruction of MG occur relatively independently and together contribute to a vicious cycle in the development of MGD.
In this review, based on the latest experimental evidence, we summarizing the pathological mechanisms about different clinical characteristics of MGD, attempting to elucidate the interaction between MG ductal obstruction and glandular dropout, aiming to better guide clinicians to diagnose and treat MGD.
Suzuki divided the menstrual cycle into six phases (phase I-VI), the straight-chain saturated fatty acids (SFA) significantly increased and the ratio of mono-unsaturated fatty acids (UFA) significantly decreased in phase II and returned to normal levels in phase III-V [31]. Meibum SFA/UFA of the androgen deficiency patients also had significant changes, showing as imbalanced content of cholesterol esters and wax esters [14]. It has known that knockout of the rate-limiting enzyme stearoyl-CoA desaturase 1 for the monounsaturated fatty acids synthesis [32] and the elongase of very long chain fatty acid (ELOVL) for very-long-chain fatty acids synthesis [33], all significantly altered SFA/UFA ratio in meibum, which associated with MG ductal epithelium hyperplasia, central ducts narrowing, and obstruction. These indicate that sex hormones fluctuation may induce MG obstruction by altering the proportion of meibum SFA and UFA [31]. Changes of SFA/UFA ration would enhance the lipid-lipid interaction strength, which makes meibum exhibit higher phase transition temperature and unsaturation, causing it transited from a relatively disordered liquid state to a more ordered solid state [34-35]. Solid meibum often secrets stagnation, gradually accumulates in the glandular duct causing MG ductal keratinization and obstruction [3, 25]. Moreover, solid meibum has a reduced fluidity makes it more difficult to spread on the tear film (TF), formatting inefficient and thinner TFFL [35], inducing a desiccative and hypertonic ocular environment, which further leads to meibocyte differentiation and meibum metabolisms abnormalities [21].
Recently, multiple targets were demonstrated to be involved in the meibum metabolisms, the most important one is peroxisome proliferator-activated receptor gamma (PPARγ). PPARγ agonist rosiglitazone upregulated meibocyte differentiation-related proteins, like adipophilin (ADFP), elongation of ELOVL4, and fatty acid binding protein 4 (FABP4) [36]. The positive effects of PPARγ on MG involved the regulation of Wnt and Hedgehog pathways [36-37], and inhibiting NF-κB pathway to correct the metabolic disorder of meibum lipids and MG obstruction in APOE deficient mice [25]. Activated NOTCH signaling suppressed Human meibomian gland epithelial cells (HMGECs) proliferation but promote its meibum synthesis [38]. In addition, 1nM insulin-like growth factor 1 (IGF-1) treatment activated PI3K/AKT and forkhead box O1 (FoxO1) pathways, simultaneously facilitated meibum production and proliferation of HMGECs [39].
One of the possible explanations is the change of eyelid tension determined by different anatomy [49-50]. In the Caucasian eyelid, levator aponeurosis penetrates the orbital septum at a higher location which induces the upper palpebral crease [49-50]. On the contrary, the lower penetrated position in Asian eyelid making the tension of eyelid increased [49]. The larger exposed ocular surface area makes both Asian pediatric and young populations showed higher PBR and more susceptible to suffer MG obstruction [51-52].
Another interpretation is the different consumption of supplements between Asian and Caucasian populations, such as fish oil, a kind of supplement contains abundant omega-3 (ω-3) fatty acids [53]. Fish oil and ω-3 are the third most popular dietary supplementations among the Caucasian population, with a frequency of 9.8%-12% [53-54], while that in the Asian population is no more than 5% [55]. It has been demonstrated that reasonable ω-3 supplementation decreased the prevalence of MGD and improved meibum quality in postmenopausal women [56]. ω-3 regulated the content of SFA [57] in meibum by altering the differentiation of HMGECs [58]. Besides, a random clinical trial demonstrated that 1.5g/d ω-3 intake lowered MGD-linked lid inflammation [59]. Through increasing the expression of resolvin D1 and cutting the production of cyclooxygenase-2, ω-3 exerted anti-inflammatory effects to inhibit cytokine activity of HMGECs [60]. These indicate that ω-3 exerts meibum-regulation and anti-inflammatory effects to refrain MG ductal hyperkeratinization and subsequent obstruction.
The development of MGD in daily CL wearers possesses obvious time regularity. The damage of CL to MG tended to concentrate on the first 2-3 years and remain stable for more than 7 years [67-68]. Compared to healthy control or long-term CL wearers (≥3 years), the related MG parameters were significantly worse in short experience (<3 years) [67-68]. This may relate to the constant inflammatory stimulation at the early stage, for even after wearing CL for several hours the expression of tear IL-17A tripled [65]. Besides, the tearing IL-6 level of CL wearers was significantly upregulated and became to normal levels after 6 days of cessation but returned to the original levels within 24h after re-wearing CL [66]. Therefore, early treatment is necessary for better control of MGD progression in CL wearers, however, why CL-related MGD lacks aggravation in the later period is still inconclusive, prospective longitudinal studies needed to be carried to obtain a reasonable explanation.
The inhibition of PPARγ and FGFR pathways may the mechanisms of age-related MG dropout, for transcriptome analysis of MG showed these two pathways were both significantly suppressed in mice aged 2 years when compared to mice aged 3 months [85]. The location of MG acinar is consistent with that of PPARγ [80], so significant acinar loss downregulated PPARγ signaling [19], intervened with meibocyte differentiation and meibum synthesis [84], which could induce ocular desiccative stress and further aggressive the deletion of progenitor and glandular dropout [21, 86]. FGFR-2 deficiency mouse showed obvious MG dropout and fewer proliferative MG progenitor [87], while exogenous supplement of fibroblast growth factor (FGF) facilitated the clonal growth of rabbit MG progenitor [88].
The toxic effects of AMs and the preservative it contained result in MG dropout by directly killing MG acinar cells [96-97]. Compared to preservative-free prostaglandin analogue (PGA), preserved PGA induced a severe MG dropout [93]. This because BAK, a preservative commonly exists in AMs eyedrops, arose atrophy and death of HMGECs even in the concentrations below the product safety requirements [96]. The toxic effects of BAK to MG are magnified when combined with PGA [94, 98]. Some non-PGA AMs, like pilocarpine and timolol, also inhibited the survival of HMGECs even at a normal dosage [99]. BAK- and AMs-induced the downregulation of AKT phosphorylation of HMGECs may the related mechanism [18, 96-97].
Except for timely removal of predisposing factors, early-stage interventions are crucial to break such vicious circle of ductal obstruction and glandular dropout. Though Meibography is often unable to detect MG dropout at an early phase of MG obstruction [116], but carrying out treatments like intense pulsed light (IPL) [117], warming compress [118], intraductal MG probing [119], and anti-inflammation is effective for preventing the ocular functional parameters deterioration and secondary acinar dropout induced by long-term MG obstruction. As for cases with significant loss of acinar, PEDF treatment or mechanical intraductal stimulation could activate progenitor to restore the growth and secretion of functional acinar, improving meibum expressibilities and TBUT [81, 119]
(Ⅱ) Administrative support: Yehong Zhuo and Yiqing Li
(Ⅲ) Provision of study materials or patients: Jiahui Tang, Zhe Liu, and Caiqing Wu
(Ⅳ) Collection and assembly of data: Siting Wu
(Ⅴ) Data analysis and interpretation: Canying Liu
(Ⅵ) Manuscript writing: All authors
(Ⅶ) Final approval of manuscript: All authors
1. Introduction
As the largest sebaceous gland in humans, the MG is situated in the upper and lower eyelids, comprising acinar and ductal components. [1] The MG produces meibum via holocrine secretion, whereby lipid-rich mature differentiated acinar cells (meibocytes) migrate, degenerate, and disintegrate within the terminal duct to release meibum, formating the tear film lipid layer (TFLL), thereby producing smooth optical surface and reducing tear evaporative rate [2]. Meibomian gland dysfunction (MGD) is characterized by ductal obstruction or glandular dropout accompanied by qualitative or quantitative abnormalities in meibum [3]. Ductal obstruction in MG is characterized by the blockage of the terminal duct due to condensed meibum of inferior quality and expressibility [4]. Glandular dropout involves functional acinar loss [5], using in vivo confocal microscopy (IVCM) can detect a significant reduction in MG acinar unit diameters and density [6]. MGD destroys tear film structure and increases tearing osmolarity, contributing to the occurrence of evaporative dry eye disease (EDED). The global prevalence of MGD ranged from 30.5%-68% [7-8], which is especially common among the elderly and has become a very common eye disease affecting life quality worldwide [9-10].The pathogenesis of MGD remains elusive. Previous hypothesis supported that the initial step of MGD involved gland obstruction due to hyperkeratinization of the ductal epithelium, leading to duct dilation and chronic intraglandular high pressure, ultimately causing secondary glandular dropout [3, 11]. However, this theory failed to fully account for certain observations in MGD patients, such as those with age-related MGD, who displayed significant glandular dropout with minimal evidence of obstruction [12]. It was known that the risk factors for MGD related to meibum metabolism disorder or damage to the MG ductal epithelial cells include chronic inflammatory infiltration and fluctuations in sex hormones [13-15]. These factors can lead to ducts hyperkeratinization and imbalance in meibum lipid composition, resulting in intraductal meibum stagnation, thereby inducing MG blockage, such prolonged blockage further increased intraductal pressure, ultimately induced functional acinar atrophy [16-17]. However, under the influence of aging and benzalkonium (BAK) chloride exposure [18-19], significant loss of progenitor cells and acinar cells can occur independently without apparent MG obstruction. Characterizing MG dropout solely as a progressive result of MG ductal obstruction overlooks the early diagnostic potential of glandular dropout, which exhibits high sensitivity (96.7%) and specificity (85%) in identifying MGD [20]. Notably, the loss of MG acini also affected meibocyte differentiation and meibum metabolism, increasing the risk of MG obstruction [21-22]. These findings indicate that glandular dropout and ductal obstruction of MG occur relatively independently and together contribute to a vicious cycle in the development of MGD.
In this review, based on the latest experimental evidence, we summarizing the pathological mechanisms about different clinical characteristics of MGD, attempting to elucidate the interaction between MG ductal obstruction and glandular dropout, aiming to better guide clinicians to diagnose and treat MGD.
2. How do risk factors determine ductal obstruction?
2.1 Chronic inflammation
IVCM detection found that the number of inflammatory cells within the MG glandular duct among MGD patients was twice that of healthy control [23]. Immunofluorescent staining of human eyelid specimens also detected CD45+ leukocyte infiltrated around MG was significantly associated with MG obstruction (r =0.414, P <0.05) [19]. The recruitment of Th17-mediated neutrophils orchestrated by the ocular immune system within the MG microenvironment may contribute to the pathogenesis of MG ductal alterations. Evidence from animal studies confirmed this theory, inflammatory model of allergic eye disease in mice showed obvious obstructive MG, central duct dilatation, and epithelium thickening, which was significantly associated with periglandular infiltration of polymorphonuclear neutrophils [24]. Besides, inflammatory stress activated by NF-κB made apolipoprotein E (APOE) deficient mice showed dilated MG duct and MG orifice plugging with increased keratinization and neutrophil infiltration [25]. Results from the organotypic culture of MG first revealed the potential mechanism, 50ng/ml neutrophil chemoattractant IL-1β exposure caused overexpression of Keratin 1 in MG ducts [13], indicating that keratinization of MG duct caused by chronically inflammatory stimulation seems to be the origination of MG obstruction.2.2 Sex hormones fluctuation
Estrogen fluctuation is a risk factor for inducing obstructive MGD, whereas androgen shows protective effects. Both androgen receptor and estrogen receptor have been reported to express in human MG [26], and common signature genes of meibum metabolisms showed a similar fashion of male and female populations [27]. However, androgen supplementation enhances the expression of MG keratinization inhibition and meibum synthesis associated genes, such as sterol regulatory element binding proteins 1 and 2 [28-29]. Compared to normal people, chronic antiandrogen medication users and complete androgen insensitivity syndrome patients showed a higher incidence of MGD with severe MG obstruction, decreased meibum quality [14], and an over two fold increased of dry eye symptoms including the sensations of dryness, pain, and light sensitivity [17]. On the contrast, extra estrogen upregulated the expression of meibum decomposition and MG keratinization associated genes [30]. Higher serum estrogen level in postmenopausal women characterized as a significant predictor of worse MG obstruction [15].Suzuki divided the menstrual cycle into six phases (phase I-VI), the straight-chain saturated fatty acids (SFA) significantly increased and the ratio of mono-unsaturated fatty acids (UFA) significantly decreased in phase II and returned to normal levels in phase III-V [31]. Meibum SFA/UFA of the androgen deficiency patients also had significant changes, showing as imbalanced content of cholesterol esters and wax esters [14]. It has known that knockout of the rate-limiting enzyme stearoyl-CoA desaturase 1 for the monounsaturated fatty acids synthesis [32] and the elongase of very long chain fatty acid (ELOVL) for very-long-chain fatty acids synthesis [33], all significantly altered SFA/UFA ratio in meibum, which associated with MG ductal epithelium hyperplasia, central ducts narrowing, and obstruction. These indicate that sex hormones fluctuation may induce MG obstruction by altering the proportion of meibum SFA and UFA [31]. Changes of SFA/UFA ration would enhance the lipid-lipid interaction strength, which makes meibum exhibit higher phase transition temperature and unsaturation, causing it transited from a relatively disordered liquid state to a more ordered solid state [34-35]. Solid meibum often secrets stagnation, gradually accumulates in the glandular duct causing MG ductal keratinization and obstruction [3, 25]. Moreover, solid meibum has a reduced fluidity makes it more difficult to spread on the tear film (TF), formatting inefficient and thinner TFFL [35], inducing a desiccative and hypertonic ocular environment, which further leads to meibocyte differentiation and meibum metabolisms abnormalities [21].
Recently, multiple targets were demonstrated to be involved in the meibum metabolisms, the most important one is peroxisome proliferator-activated receptor gamma (PPARγ). PPARγ agonist rosiglitazone upregulated meibocyte differentiation-related proteins, like adipophilin (ADFP), elongation of ELOVL4, and fatty acid binding protein 4 (FABP4) [36]. The positive effects of PPARγ on MG involved the regulation of Wnt and Hedgehog pathways [36-37], and inhibiting NF-κB pathway to correct the metabolic disorder of meibum lipids and MG obstruction in APOE deficient mice [25]. Activated NOTCH signaling suppressed Human meibomian gland epithelial cells (HMGECs) proliferation but promote its meibum synthesis [38]. In addition, 1nM insulin-like growth factor 1 (IGF-1) treatment activated PI3K/AKT and forkhead box O1 (FoxO1) pathways, simultaneously facilitated meibum production and proliferation of HMGECs [39].
2.3 Incomplete blinking
Ruled by cranial nerve seven (CN VII), the human blinking process identified as having four phases: downstroke, turning point, upstroke, and the interblink interval [40]. During the phase of downstroke to upstroke, blinking facilitates the spread and distribution of TF, and the formation of TFLL by continuous meibum secretion regulated by synchronized eyelid muscular actions [3], while the evaporation and break up of TF arise in the interblink interval. Abnormal blinking is characterized as incomplete blinking (IB) or higher partial blinking rate (PBR) which is associated with MG obstruction, worse meibum quality, TBUT, and OSDI scores [41]. A week blinking training could significantly improve the MGD-related symptoms of IB populations [42]. A plausible mechanism is that IB inhibits the squeezing power of orbicularis muscle and Riolan muscle, leading to meibum excretion reduction and stasis in the glandular ducts which finally caused MG obstruction [3]. IB also arises TFLL unevenly distributed [41], causing increased tear evaporation rate and a desiccative ocular environment [43], which in turn leads to MG differentiation and meibum production abnormalities, exacerbating MG obstruction [21]. Patients with secondary IB diseases, like CN VII palsy, thyroid eye disease (TED), and reduced corneal sensitivity caused by diabetic peripheral neuropathy also showed similar obstructive MGD features [44-46].2.4 Racial difference
The difference between the human race is quite an interesting risk factor of MGD. Compared to Western countries, age-matched people in Asian countries exhibit a higher incidence of MGD [7, 47]. After excluding the difference of diagnostic criteria and environment, this phenomenon remained the same that adult Asian descendants appeared poorer meibum quality, OSDI scores, TF stability, TFLL structure, and tear osmolarity [7, 47- 48].One of the possible explanations is the change of eyelid tension determined by different anatomy [49-50]. In the Caucasian eyelid, levator aponeurosis penetrates the orbital septum at a higher location which induces the upper palpebral crease [49-50]. On the contrary, the lower penetrated position in Asian eyelid making the tension of eyelid increased [49]. The larger exposed ocular surface area makes both Asian pediatric and young populations showed higher PBR and more susceptible to suffer MG obstruction [51-52].
Another interpretation is the different consumption of supplements between Asian and Caucasian populations, such as fish oil, a kind of supplement contains abundant omega-3 (ω-3) fatty acids [53]. Fish oil and ω-3 are the third most popular dietary supplementations among the Caucasian population, with a frequency of 9.8%-12% [53-54], while that in the Asian population is no more than 5% [55]. It has been demonstrated that reasonable ω-3 supplementation decreased the prevalence of MGD and improved meibum quality in postmenopausal women [56]. ω-3 regulated the content of SFA [57] in meibum by altering the differentiation of HMGECs [58]. Besides, a random clinical trial demonstrated that 1.5g/d ω-3 intake lowered MGD-linked lid inflammation [59]. Through increasing the expression of resolvin D1 and cutting the production of cyclooxygenase-2, ω-3 exerted anti-inflammatory effects to inhibit cytokine activity of HMGECs [60]. These indicate that ω-3 exerts meibum-regulation and anti-inflammatory effects to refrain MG ductal hyperkeratinization and subsequent obstruction.
2.5 Contact lens
Whatever soft or rigid lens, daily contact lens (CL) wearing is recognized as a risk factor of MGD [61-62]. Daily CL wearers showed obvious MG obstruction with meibum abnormalities, OSDI and CFS were also significantly higher [62]. Two explanations may help to describe this phenomenon. Mechanical trauma from the CL induced desquamated ductal epithelial cells accumulated in the opening end of the duct [63], the continuous secretion of meibum eventually blocked MG [61-62, 64]. Besides, CL wearing also increased the expression level of IL-17A, TNF-α, and IL-6 in tears [65-66], which made MG face the risk of ductal hyperkeratinization and blockage.The development of MGD in daily CL wearers possesses obvious time regularity. The damage of CL to MG tended to concentrate on the first 2-3 years and remain stable for more than 7 years [67-68]. Compared to healthy control or long-term CL wearers (≥3 years), the related MG parameters were significantly worse in short experience (<3 years) [67-68]. This may relate to the constant inflammatory stimulation at the early stage, for even after wearing CL for several hours the expression of tear IL-17A tripled [65]. Besides, the tearing IL-6 level of CL wearers was significantly upregulated and became to normal levels after 6 days of cessation but returned to the original levels within 24h after re-wearing CL [66]. Therefore, early treatment is necessary for better control of MGD progression in CL wearers, however, why CL-related MGD lacks aggravation in the later period is still inconclusive, prospective longitudinal studies needed to be carried to obtain a reasonable explanation.
2.6 Demodex infestation
The found of Demodex in the obstructive and dilatated ducts with abnormal keratinization of MGD patients suggested that ocular Demodex infestation may be a potential risk factor of MG obstruction [69]. Demodex folliculorum (D.f) and Demodex brevis (D.b) are the only two types of mites that parasitic to humans [70]. Clinical observations found that patients with ocular Demodex infestation showed higher incidence and degree of MGD than healthy populations [71]. MG secretions and eyelashes from ocular Demodex infection positive patients showed complicated bacteria profiles (higher proportion of P. acnes, S.aureus, Staphylococcus, and Streptococci) [72-73]. Demodex’s intestine also harboured Bacillus oleronius which could trigger a host immune response via producing pro-inflammatory proteins [74]. This indicated that Demodex may act as the vehicle of bacterias and induce host inflammatory response to increase the risk of MG obstruction [73-74]. Our former study also found almost 90% of young ocular demodicosis patients possessed MGD, which was significantly associated with D.b infestation rather than D.f [71]. Besides, D.b infestation was associated with worse meibum quality [75]. This was consistent with the fact that D.b tends to the resident in MG [76], while D.f prefers parasitizing in the lash follicle [77], the mechanical irritation of D.b may lead to reactional MG epithelial hyperplasia and glandular obstruction [77]. The chitinous exoskeleton of D.b may also act as a foreign body and cause granulomatous reactions [76]. Therefore, a higher Demodex infection rate, particularly D.b, may one of the risk factors of MG obstruction.3. How do risk factors determine glandular dropout?
3.1 Aging
MGD is considered an age-related disease [7, 47]. Obvious functional and morphologic changes of MG can be detected in asymptomatic populations beyond 35 years-old [47]. With aging, the dropout ratio of MG (Meiboscore) raised [5], while the MG acinar unit diameters and densities are significantly decreased [6]. How aging affects MGD is quite disputable. Previous studies suggested that aging firstly triggered ductal hyperkeratosis and obstruction to result in MG atrophy and dropout [3]. However, recent research on MG cadaver specimens from the aged person showed that there is a significant loss of progenitor cells but without ductal hyperkeratosis [78]. IVCM detection showed that human aging MG exhibited signs of atrophic but nonobstructive [6]. Similarly, proliferative progenitor cells were largely absent in age‐related MGD mouse model [79], but hyperkeratinization of MG was failed to detect [78-80]. Compared to young mice, the expression of Ki67, which indicates the proliferation activity of cells, was decreased in MG of elder mice [80]. Subconjunctival injection of pigment epithelium-derived factor (PEDF), a stem cell proliferation promoter, increased the expression of stem cell markers ΔNp63 and Lrig1 in acinar basal progenitor cells, restored MG acinar area, and improved the TBUT of age-related MGD mice [81]. Therefore, a probable mechanism of age-related MGD is the depletion of progenitor cells which leads to MG renewal decompensation, arising subsequent MG atrophy, and dropout [19]. Recent in vivo and in vitro models have substantiated this hypothesis that cell senescence induced by high-glucose stimulation markedly induced the depletion of progenitor cells in MG, leading to significant functional acinar dropout [82-83]. However, the hyperkeratinization and obstruction of aged MG may attribute to other risk factors [78, 84].The inhibition of PPARγ and FGFR pathways may the mechanisms of age-related MG dropout, for transcriptome analysis of MG showed these two pathways were both significantly suppressed in mice aged 2 years when compared to mice aged 3 months [85]. The location of MG acinar is consistent with that of PPARγ [80], so significant acinar loss downregulated PPARγ signaling [19], intervened with meibocyte differentiation and meibum synthesis [84], which could induce ocular desiccative stress and further aggressive the deletion of progenitor and glandular dropout [21, 86]. FGFR-2 deficiency mouse showed obvious MG dropout and fewer proliferative MG progenitor [87], while exogenous supplement of fibroblast growth factor (FGF) facilitated the clonal growth of rabbit MG progenitor [88].
3.2 Topical anti-glaucoma medications
Chronic use of anti-glaucoma medications (AMs) is a risk factor of MGD. Compared to the normal population, the incidence of MGD among patients who received AMs for more than 1 year is as high as 80% [89-90]. The burden of antiglaucoma (BAG), which represents the duration [89], frequency [91], compliance [92], and drug number of AMs [93-94], is significantly correlated with the severe degree of MGD [91]. Accompanied by worse scores of OSDI and TBUT [95], high-BAG population showed increased MG dropout, lower MG acinar density, and thinner TFLL [91, 93, 94].The toxic effects of AMs and the preservative it contained result in MG dropout by directly killing MG acinar cells [96-97]. Compared to preservative-free prostaglandin analogue (PGA), preserved PGA induced a severe MG dropout [93]. This because BAK, a preservative commonly exists in AMs eyedrops, arose atrophy and death of HMGECs even in the concentrations below the product safety requirements [96]. The toxic effects of BAK to MG are magnified when combined with PGA [94, 98]. Some non-PGA AMs, like pilocarpine and timolol, also inhibited the survival of HMGECs even at a normal dosage [99]. BAK- and AMs-induced the downregulation of AKT phosphorylation of HMGECs may the related mechanism [18, 96-97].
3.3 Chemotherapy
Chemotherapeutic drugs induced a higher prevalence of MGD, such as TS-1 [100] and aromatase inhibitors [101], and of which the adverse events of MG induced by anticancer TS-1® combination capsules are most common. Containing tegafur (FT) and 5-fluorouracil (5-FU), TS-1® inhibits cellular DNA synthesis and is widely used to treat digestive system cancer [102]. Matsumoto first reported that 10 patients all showed foreign body sensation and visual discomfort with significant MG atrophy and dropout after receiving TS-1 chemotherapy for more than 4 months [100]. TS-1 chemotherapy reduced MG acinar area in a dose-dependent manner, especially during the period of 3-6 months after chemotherapy [102]. FT and 5-FU in tears during TS-1 treatment reached (1.94 ± 0.71) and (0.17 ± 0.11)μ g/mL [103]. Chronically exposed to this concentration significantly suppressed the proliferation of various human ocular epithelial cells (cornea, conjunctiva, and retina) [104], suggesting that chemotherapeutic drugs accumulated in tears may show extensive toxicity to inhibit the survival of MG acinar cells, arising severe glandular atrophy and dropout in a short time [105]. Unfortunately, research involving animal models of systemic chemotherapy tend to pay little attention to the damage of MG or ocular surface caused by chemotherapeutic drugs, which needs to be concerned in the future.3.4 Radiotherapy
Periocular radiotherapy (PRT), as a typical treatment to periorbital tumors [106], is found to cause MG dropout [107]. PRT ranged from 30-70 Gy induced MG dropout in a radiation dose-dependent manner [108] with no trails of ductal dilatation or obstruction [109]. Even in the lowest radiation dose (≤30 Gy), PRT-treated patients also possessed significant MG dropout, and the scores of OSDI, TBUT, and CFS were all significantly deteriorated [107-109]. Besides, among patients only accepted unilateral PRT treatment, their bilateral upper and lower MG both showed severe MG dropout [108]. PRT-induced MG dropout seems to be irreversible, the glandular dropout of patients still in a severe state after discontinued PRT for 6 months [109]. Similar to the sebaceous gland and epidermal stem cells, MG also have activated progenitor [19], which may make it vulnerable to radiation-induced DNA damage. Histopathological results from patients who had received PRT detected obvious atrophy of MG acinar [110], which further indicated that the direct damage of PRT to MG acinar and progenitor cells may the potential pathogenic mechanism.4. The vicious cycle of MG ductal obstruction and glandular dropout
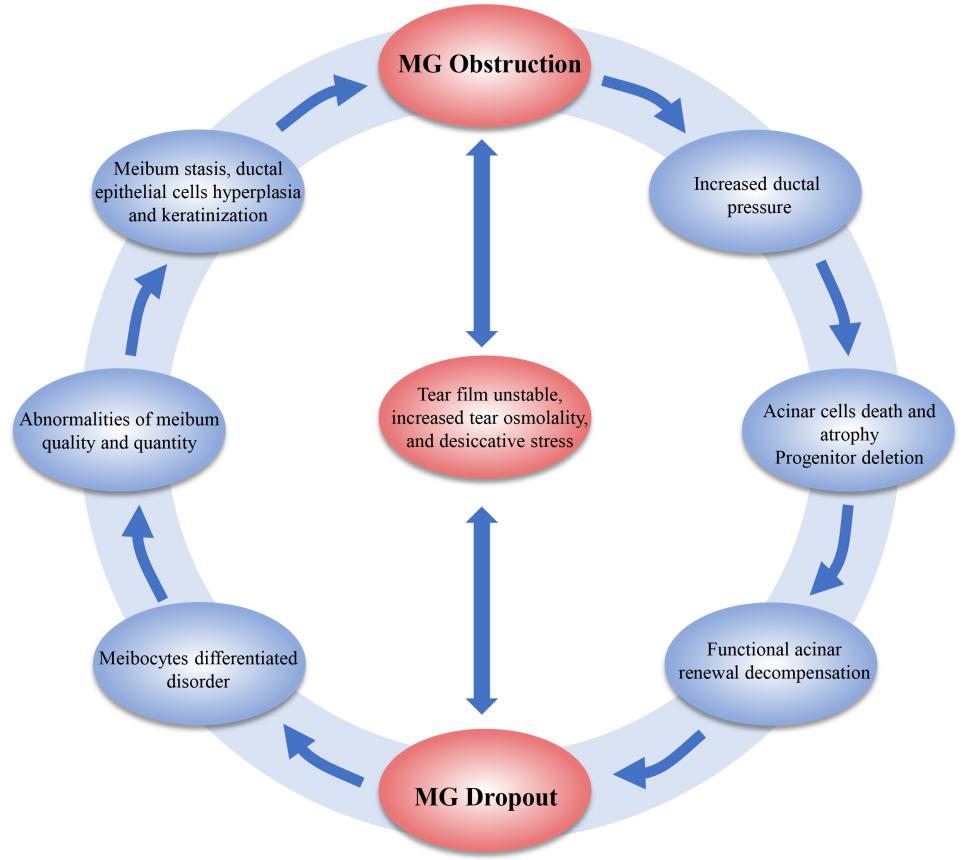
The role of MG obstruction and MG dropout are interrelated, Meiboscore correlates with meibum expressibility (r = 0.53, P < 0.001) [111], compared to Meiboscore 0, patients of Meiboscore 1-3 showed more frequent MG obstruction [20]. Due to the risk factors of ductal obstruction and glandular dropout are not exactly identical, they occur in sequence, but together constitute a vicious circle of MGD pathogenesis. MG obstruction induced increased glandular pressure which gradually appeared progenitor cell renewal decompensation, functional acinar atrophy and dropout [1, 3, 112]. PPARγ is co-located with glandular acinar [19, 80], MG dropout made the PPARγ signaling suppressed [22, 25] and the number of acinar cells differentiate into meibocyte decreased [19, 80] which resulted in meibum SFA/UFA and lipids-protein abnormalities [84], increased hardness and stickiness, but decreased fluidity, causing meibum secretory stagnation [34-35], ductal keratinization, and subsequent obstruction [3, 25]. What’s more, both ductal obstruction and glandular dropout would destruct TFLL structure, inducing ocular desiccative stress and tear evaporation accelerated [113-115], which in turn intervene with meibocyte differentiation and progenitor cell proliferation and renewal, aggravating MG obstruction/dropout (Figure 1) [21].Figure 1 The vicious circle of MG ductal obstruction and glandular dropout
Except for timely removal of predisposing factors, early-stage interventions are crucial to break such vicious circle of ductal obstruction and glandular dropout. Though Meibography is often unable to detect MG dropout at an early phase of MG obstruction [116], but carrying out treatments like intense pulsed light (IPL) [117], warming compress [118], intraductal MG probing [119], and anti-inflammation is effective for preventing the ocular functional parameters deterioration and secondary acinar dropout induced by long-term MG obstruction. As for cases with significant loss of acinar, PEDF treatment or mechanical intraductal stimulation could activate progenitor to restore the growth and secretion of functional acinar, improving meibum expressibilities and TBUT [81, 119]
5. Conclusion and outlooks
Comprehensive analysis in the diagnosis and treatment of MGD is necessary since the occurrence of MG obstruction and MG dropout is relatively independent with a certain sequence, which largely depends on the risk factors patients suffered. MG ductal obstruction and glandular dropout interact with each other, long-term obstruction induces acinar cells atrophy and progenitor cells deletion, whereas glandular dropout aggravates meibocyte differentiation and meibum synthesis, increasing the risk of ductal blockage and MGD parameters deterioration. Therefore, timely intervention and removal of predisposing factors are quite important for avoiding falling into the vicious circle of MGD. The construction of HMGECs and MG animal models reveal some reliable molecular basis, PPARγ, FGFR, PI3K-AKT, and NF-κB pathways play an essential role in controlling the proliferation, differentiation, and meibum lipids metabolism of MG which provides potential treatment targets for clinicians. However, the current experimental evidence is still inadequate, the pathogenic mechanisms of some risk factors such as Demodex infestation remain unknown. More sophisticate animal and cellular studies should carry out to clarify the concrete components of meibum lipids with pathogenicity and the process of keratinization of MG ductal epithelial cells. Additionally, effective therapies and potential molecular targets for restoring MG functional acinar also need to be further investigated and validated.Correction notice
NoneAcknowledgement
NoneAuthor Contributions
(Ⅰ) Conception and design: Qi Zhang and Yiqing Li(Ⅱ) Administrative support: Yehong Zhuo and Yiqing Li
(Ⅲ) Provision of study materials or patients: Jiahui Tang, Zhe Liu, and Caiqing Wu
(Ⅳ) Collection and assembly of data: Siting Wu
(Ⅴ) Data analysis and interpretation: Canying Liu
(Ⅵ) Manuscript writing: All authors
(Ⅶ) Final approval of manuscript: All authors
Funding
This work was supported by the National Natural Science Foundation of China (82371021, 82201142), the Science Foundation of Guangdong Province (2023A1515012336, 2023A1515010091, 2023A1515030238).